谁能想到,让我们记住那些美好童年回忆和复杂的数学公式的竟然是一种类似于"发炎"的过程和 DNA 损伤吗?期刊 Nature 上最近发表的一项研究表明,长期记忆储存依赖于"大脑炎症"和 DNA 损伤。是的,你没有看错!这种应对逆境的生物反应正是锻造我们记忆力的关键工具。这两个看似不合群的角色——神经"发炎"和 DNA 损伤是如何共同塑造我们的长期记忆存储的呢?快来随小 M 一起来看看吧~

听到 “发炎” 和 “DNA 损伤”,可能首先想到的是疾病或损伤,但在我们的大脑中,这两个过程却是形成长期记忆的关键步骤,与我们大脑中一种名为海马神经元的特殊细胞有关。
这些神经元如同建筑师一般,将各种信息组装成微电路,存储我们生活中的各种瞬间。然而,这个过程并非毫无 “代价”,大脑中的海马神经元会经历一个能量密集的过程,此时神经元的 DNA 双链会暂时发生断裂 (DNA double strand breaks, DSBs),并涉及一种叫做 TLR9 信号分子的激活。这个过程就像是神经元的一次小型“发炎”,为记忆储存打下了坚实的基础[1]。DNA 双链断裂与 DNA 修复被认为是神经可塑性 (Neuroplasticity) 的基础,神经可塑性对于学习和记忆等认知过程以及大脑发育、感觉训练和大脑损伤的恢复至关重要[2]。

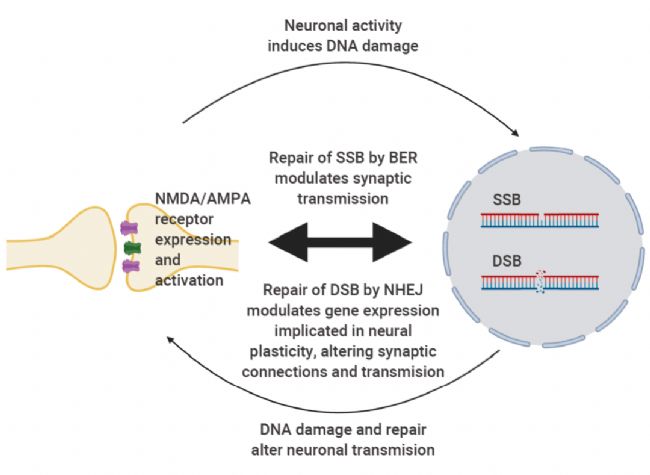
图 1. 神经可塑性与 DNA 损伤和修复之间相互作用的简化模型[2]。
突触处 NMDA/AMPA 受体的激活会诱导 SSB 或 DSB,并分别通过 BER 或 NHEJ 促进其修复。反之,DNA 损伤和修复会改变这些受体的表达和活性,从而调节神经元基因表达,导致可塑性发生变化。SSB:单链 DNA 断裂; DSB: 双链 DNA 断裂。BER: 碱基切除修复; NHEJ: 非同源末端连接。

近期,发表于 Nature 的一项研究发现,在长时间学习后,兴奋性海马 CA1 神经元出现了双链 DNA (dsDNA) 断裂、核膜破裂以及释放组蛋白和 dsDNA 片段等一系列现象。这些早期事件之后,一些神经元开始表现出炎症表型,涉及到 TLR9 的信号通道的激活,同时也出现了 DNA 损伤修复复合物的积累。如果 TLR9 功能受损,基本的记忆机制可能会变为基因组不稳定和认知障碍的开端,与加速衰老、精神疾病和神经退行性疾病有关。
TLR9 参与的关键作用:负责 DNA 的损伤修复,参与纤毛的生成和周围神经元网的建立。
▐ 免疫应答基因 (TLR9) 在记忆形成中的关键作用作者首先对背侧海马神经元的转录谱进行了深入分析,发现在情境恐惧调节(Contextual fear conditioning, CFC) 后 96 小时或 21 天内,基因表达谱与远程记忆的差异显著。研究人员观察到大多数差异表达基因是参与核酸感应和细胞因子释放的免疫应答基因 (图 2A)。
在免疫应答基因中,TLR9 及其下游 NF-κB 信号通路突出,并观察到 TLR9 基因和蛋白水平的上调,以及 TLR9 与晚期内体 (Endosomes) 和溶酶体标志物 LAMP2 的共定位增加 (图 2B),说明 TLR9 向内体的运输增强,有助于 DNA 识别和 NF-κB 激活。这些发现揭示了免疫应答基因在记忆形成中的关键作用。

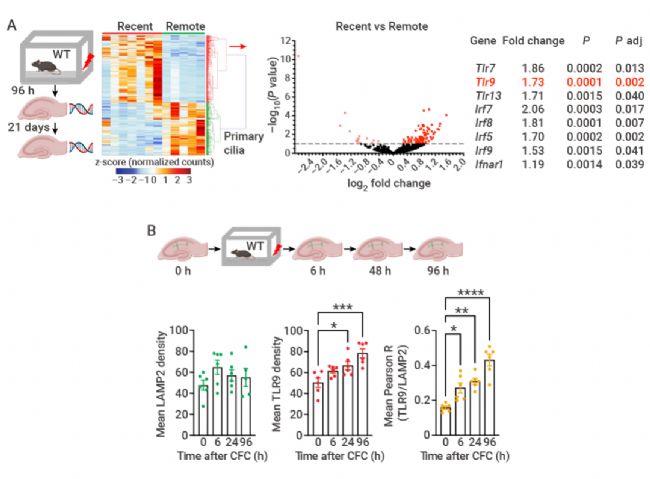
图 2. CFC 后的基因表达谱图及 TLR9 蛋白水平,TLR9/LAMP2 在不同时间的共定位[3]。
A. 与 CFC 后 21 天相比,Bulk RNA-seq 显示 CFC 后 96 小时获得的小鼠海马中 441 个基因的表达增加。火山图显示与炎症和 TLR 信号传导相关的基因表达显着增加。B. TLR9 蛋白水平以及 TLR9 与 LAMP2 在 CFC 后不同时间的共定位。LAMP2 水平无波动,TLR9 水平及其与 LAMP2 的共定位在 CFC 后 6 小时增加,96 小时后达到峰值。
▐ CFC 触发 dsDNA 断裂和 DNA 损伤修复 (DDR)研究人员使用 dsDNA 断裂结合磷酸组蛋白 γH2AX 特异性抗体进行免疫荧光标记,发现 CFC 后的 1 小时和 3 小时内,部分神经元中 dsDNA 断裂的数量显著增加,并且具有神经元特异性。虽然随后核病灶的数量减少,但在单个神经元内出现了更大的 γH2AX 标记病灶 (图 3A-B),且核周 γH2AX 信号与 TLR9 信号存在共定位 (图 3C)。
这些发现表明,在一些神经元中单独的 γH2AX 和 dsDNA 或复合物的形式从细胞核中含有 TLR9 的核周位点释放出来。另一方面,研究发现 γH2AX 信号与炎症信号传导有关,如其与 RELA ( NF-κB 家族成员,在激活后易位至细胞核) 存在高度共定位。这些发现表明,记忆形成过程中的神经元活动可能会引发 DNA 损伤,并启动 DNA 修复机制。

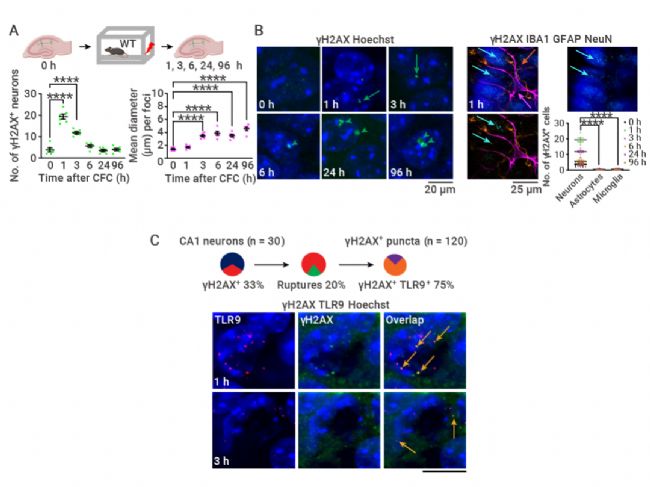
图 3. CFC 后的 DNA 损伤和 DDR (DNA 损伤修复)[3]。
A. CFC 之后 γH2AX 点状的神经元数和 γH2AX 病灶大小。B. γH2AX 相对于星形胶质细胞和小胶质细胞在神经元中的定位。C. 核外 γH2AX 与 TLR9 共定位。
▐ CA1 神经元中的 TLR9 是情景记忆所必需的炎症反应是学习-诱导的 DNA 损伤的副作用还是有助于记忆的形成?作者通过在背侧海马 CA1 区域的神经元中特异性敲除 TLR9,发现敲除后的小鼠显示出受损的情景记忆,即冻结行为 (Freezing Behaviour, 指动物对特定刺激的反应,动物会激活不同性质的防御模式,包括冻结和主动的战斗或逃跑反应。冻结是一种行为抑制形式,伴随着副交感神经主导的心率减慢) 显著减少 (图 4A),且星形胶质细胞和小胶质细胞的敲除并未影响记忆缺陷,这也与先前的发现一致。同时,TLR9 拮抗剂 ODN2088 显著损害 CFC,而 cGAS-STING 抑制剂 RU-521 和 H-151 则无效 (图 4B)。此外,TLR9 敲低也破坏了 CFC 诱导的基因表达。以上结果证明了神经元特异性 TLR9 介导的 dsDNA 传感在情境记忆的形成和持久中的作用。

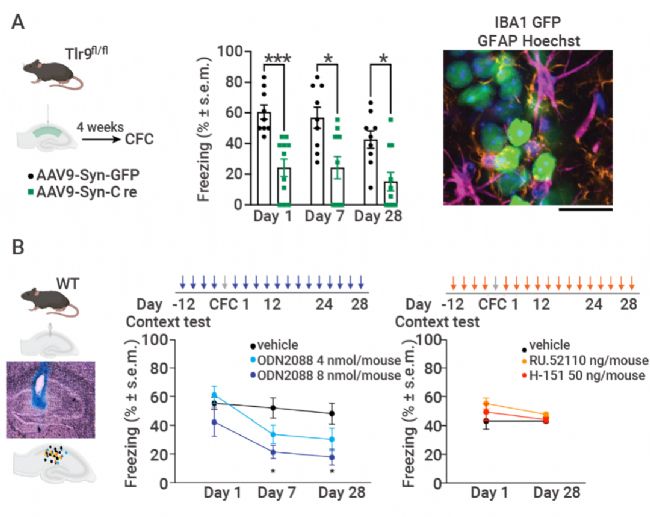
图 4. 海马 TLR9 神经元特异性缺失后情境记忆受损[3]。
A. 与注射 Syn-GFP 的对照组相比,在海马内注射 Syn-cre 的 TLR9fl/fl 小鼠的情景测试中,冻结持续减少。B. ODN2088 对 TLR9 的药理抑制作用对 CFC 造成剂量依赖性损害。
此外,作者通过实验观察 WT 和 TLR9fl/fl 小鼠海马部位,发现神经元中的 dsDNA 断裂数量显著增加。TLR9 的缺失破坏了核和中心体 DDR,使得在 CFC 时,CA1 神经元无法招募 DDR 复合物或形成纤毛和 PNNs,证实了 TLR9 在 DDR、纤毛形成和神经周围网 (Perineuronal nets, PNN) 积累中的作用。
今天小 M 和大家一起了解到,通过学习诱导的 TLR9 信号传导,神经元将 DNA 损伤与 DDR 联系起来,对记忆的稳定性和持久性起关键作用。TLR9 激活可能由 γH2AX 和 dsDNA 片段触发。TLR9 参与了 DNA 传感,而不是 CFC 中更为传统的 cGAS-STING 路径,提示神经元采用了基于免疫的记忆机制。因此,保持 TLR9 炎症信号传导的完整性成为神经认知缺陷的有前途的预防策略,或许可以为衰老、精神疾病和神经退行性等疾病提供一些新见解~

| PLX5622口服 CSF1R 抑制剂,能透过血脑屏障的、小胶质细胞清除剂。 |
| PLX5622 in AIN-76A DietPLX5622 饲料形式,1200 ppm |
| ODN 2088TLR3,TLR7 和 TLR9 抑制剂 |
| RU.521cGAS 抑制剂 |
| H-151选择性和共价的 sting 拮抗剂,可用于自身免疫性疾病的研究。 |
| γH2AX antibody用于 WB, ICC/IF |



[1] Madabhushi R, et al. Activity-Induced DNA Breaks Govern the Expression of Neuronal Early-Response Genes. Cell. 2015 Jun 18;161(7):1592-605.
[2] Konopka A, et al. The Role of DNA Damage in Neural Plasticity in Physiology and Neurodegeneration. Front Cell Neurosci. 2022 Jun 23;16:836885.
[3] Jovasevic V, et al. Formation of memory assemblies through the DNA-sensing TLR9 pathway. Nature. 2024 Apr;628(8006):145-153.


