近,来自比利时根特大学的专家利用WIWAM XY植物表型系统发表了题为Drought resistance is mediated by divergent strategies in closely related Brassicaceae的文章,发表在植物学期刊New Phytologist上。
将大规模自动化理念和工业级零件和设备整合入植物成像系统的厂家,SMO公司在植物表型成像分析领域处于的技术前列,大面积叶绿素荧光成像系统使WIWAM成为植物表型分析与功能成像领域为*仪器设备,植物生长、胁迫响应等测量参数达几百个。工业级部件品质使系统非常耐用,基本免维护,与同类产品相比,特点突出。目前WIWAM植物表型平台分为WIWAM XY,WIWAM Line以及WIWAM Conveyor 3个系列。野外移动版称为WIWAM Screening。
WIWAM XY是一款高通量可重复性表型机器人,用于对小型植物, 如拟南芥植物的研究。该机器人可定期对多种植物参数进行自动化灌溉和并测量多种植物生长参数。WIWAM XY代替了很多手工处理,省时省钱,精度*。

WIWAM XY由花盆定位桌面, 不同个体线路, 上方机器臂以及1或多个成像或称重/浇水站组成。全套系统可以安装在现有生长室,内置高品质工业部件,可按用户要求进行定制。
植物在各自花盆内生长, 预设时间间隔,机器臂提取植物, 将其带到成像和称重浇水工作站。机器臂按照程序设定移动到花盆上方,并将其提升脱离桌面。设备可配RFID读取装置以及花盆可贴上FID 标签,可作为额外花盆识别法 ,识别和校正桌面上因手工花盆安置造成的错误。通常旁边取景照相机从不同角度获得图像。成像站可安装一系列照相机系统。组合称重/浇水站集成在机器臂上。花盆中植物在浇水时旋转以获得水分布 。灌溉精度*可达 +/- 0.1 mL。另外,灌溉可基于自动目标重量计算或固定量。在整个实验过程中,可控制土壤湿度水准 。集成光,温度和湿度传感器可监控温度,详细记录实验生长条件。
植物表型机器人WIWAM XY产品特点
1、浇水时花盆旋转以获得水分布
2、高精度灌溉(达0.1mL !).
3、WIWAM XY 可配置环境传感器
4、WIWAM XY 配有直观用户界面
5、开放式数据库结构
6、可提供全定制系统、花盆数量可扩展
7、高环境适应性、耐受苛刻的调节后环境
图像分析和数据可视化
WIWAM XY有VIB开发的图像分析和数据可视化软件支持此软件包,称为 PIPPA,是中央网络界面和数据库,一方面用来为不同类型的WIWAM植物表型平台提供管理的工具,另一方面用于分析图像和数据 。
PIPPA 与该平台通讯,通过将PIPPA网络界面生成的实验结果传到平台。每个花盆的处理和基因型信息已在数据库限定以确保在整个实验中的数据一体性。实验期间,,PIPPA 对来自平台的称重,灌溉测量,环境数据,错误记录以及图像信息进行处理分析。PIPPA支持这些图像后续处理 (旋转/收获/等) 。图像分析文本可以在PIPPA 界面初始化, 可设置于网络服务器运行 (独立版本) 或计算机群运行,以快速生成结果。 随后,通过检查数据是否在特定阈值之内可在网络几面对输出文本进行验证,例如,是否生长相关性状,如植物枝条面积一段时间内是否增加。
平台收集的环境和称重/浇水数据以及图像处理数据在PIPPA 显示,图表生成组件生成图表,支持几种类型图表,可聚焦以及输出文档。数据也可从文件导入PIPPA 或从PIPPA 数据库导出数据到文档,将应用范围大化。另外,因可直接获得所有图像数据, 你也可通过喜欢的分析文本轻松处理图像。
IT 解决方案和储存
WIWAM 软件在工业计算机上运行,触摸屏 。该软件配有用户友好图形界面, 用于控制机器人站行为以及以*灵活度设计设计实验 。可同时运行多组实验,可运行不同随机模式 ,可及时规划单个植株或一组植株的处理。 在预设启动时间,PC机将向工业PLC 发送指令, 照管机器人移动 。所有成像,称重/浇水以及环境数据均可存于SQL数据库,记录后可用于分析记录 。系统采用了开放式数据库结构,可以直接获取图像。该平台可以与高性能计算相连,用于分析储存数据或者可与本地服务器设施整合。
SMS 邮件服务可以通知用户机器报警和错误,可尽快进行用户干涉。系统可于任一点暂停和停下,UPS (不间断电源)可防止数据丢失和确保在停电后全系统恢复。该软件也有平台管理员系统设置和维护行为通道。
Drought resistance is mediated by divergent strategies in closely related Brassicaceae
Summary
Droughts cause severe crop losses worldwide and climate change is projected to increase their prevalence in the future. Similar to the situation for many crops, the reference plant Arabidopsis thaliana (Ath) is considered drought-sensitive, whereas, as we demonstrate, its close relatives Arabidopsis lyrata (Aly) and Eutrema salsugineum (Esa) are drought-resistant.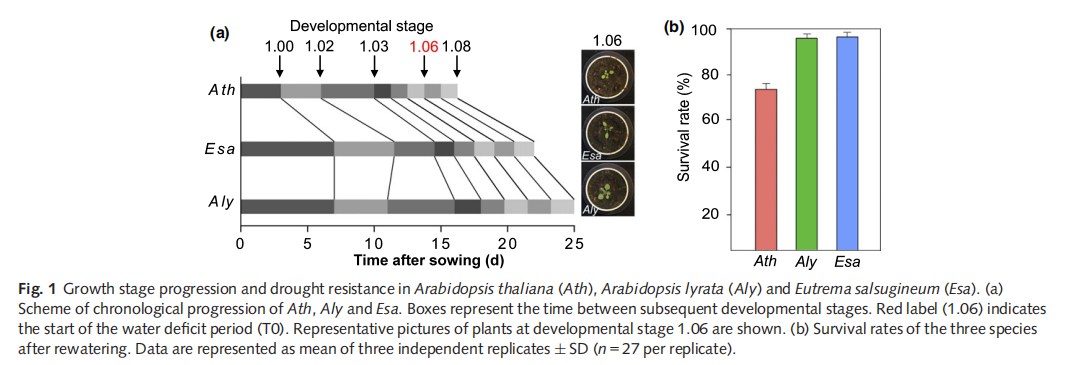
To understand the molecular basis for this plasticity we conducted a deep phenotypic, biochemical and transcriptomic comparison using developmentally matched plants.
We demonstrate that Aly responds most sensitively to decreasing water availability with early growth reduction, metabolic adaptations and signaling network rewiring. By contrast,Esa is in a constantly prepared mode as evidenced by high basal proline levels, ABA signaling transcripts and late growth responses. The stress-sensitive Ath responds later than Aly and earlier than Esa, although its responses tend to be more extreme. All species detect water scarcity with similar sensitivity; response differences are encoded in downstream signaling and esponse networks. Moreover, several signaling genes expressed at higher basal levels in both
Aly and Esa have been shown to increase water-use efficiency and drought resistance when overexpressed in Ath.
Our data demonstrate contrasting strategies of closely related Brassicaceae to achieve drought resistance
Plant growth conditions and drought treatment
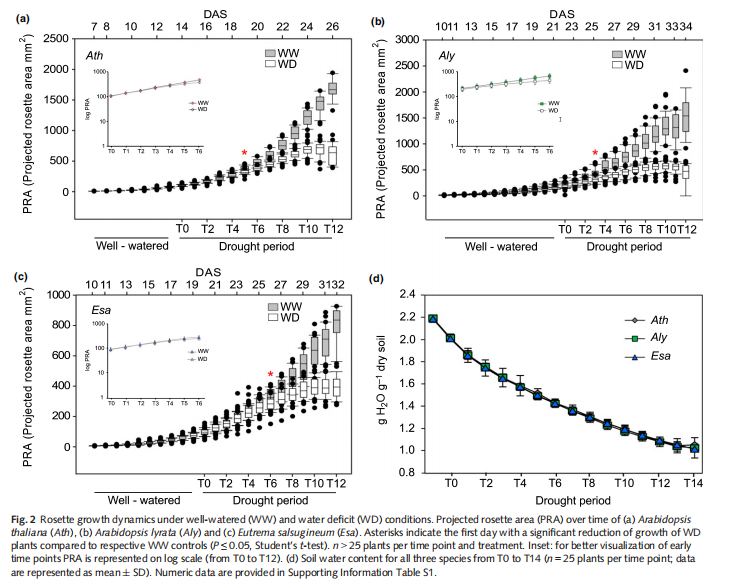
The plant phenotyping platform (WIWAM xy) at VIB Ghentwas used for high-throughput phenotypic characterization. Pots were prepared as described (Skirycz et al.,2011b). Briefly, all pots (128 per species) were radio frequencyidentification (RFID)-tagged and the dry soil weight of individual pots was calculated. Three to four plants were sown after 4 d of stratification at 4°C in the dark. Pots were placed on WIWAM xy and covered with plastic film for 3 d to maintain humidity. On day 4 the cover was removed and the well-wateredcondition (WW) of 2.19 g water g1 soil was maintained robotically. When two complete open cotyledons were observed in allpots, one average-sized seedling per pot was kept. Daily, images of the plants were taken, each pot was weighted, positions were randomized and water was added to precisely maintain WW conditions.
Water deficit (WD) treatment started when leaf 6 (L6) was initiated on the apex (1 mm, developmental stage 1.06) as judged by manual inspection (Boyes et al., 2001). Watering for the WD group (78 pots per species) was stopped at 14 (Ath), 20 (Esa) and 22 (Aly) d after sowing (DAS). After 15 d without watering, plants
were rewatered and survival was scored 3 d later. Two independent replicates were performed in trays. Plants were grown under constant environmental conditions: 16 h day, 21°C, 55% relative humidity and 110–120 lmol m2 s 1 light intensity.





