人体肠道菌群源源不断地将来自宿主和饮食的天然产物转化为生物活性代谢物。膳食脂肪是必需的微量营养素,经过脂肪分解,释放游离脂肪(fatty acids, FAs)供小肠吸收。肠道共生菌将一些不饱和脂肪酸(如亚油酸)修饰成各种肠道脂肪酸异构体,这些异构体调节宿主代谢并具有抗癌特性。然而,人们对这种饮食—微生物脂肪酸异构化如何影响宿主的粘膜免疫系统知之甚少。
近日,中科院分子细胞科学创新中心宋昕阳与美国哈佛医学院Dennis L. Kasper院士共同通讯在Nature(IF=64.8)上发表了题为“Gut microbial fatty acid isomerization modulates intraepithelial T cells”的研究论文。他们发现肠道微生物以亚油酸(Linoleic Acid, LA)等常见脂肪酸为食,并将其转化为共轭亚油酸(亚油酸的同分异构体,CLAs)。CLAs作为生物级联反应的信号,最终刺激一种特定类型的免疫细胞亚群CD4+CD8αα+上皮内淋巴细胞(CD4+CD8αα+ IELs)的发育并驻留在小肠中。而这种级联反应被阻断后,小鼠更容易被常见的食源性病原体鼠伤寒沙门氏菌所感染。
爱必信(上海)生物科技有限公司提供生化试剂 共轭亚油酸abs47051161

肠道FAs诱导CD4+CD8αα+ IELs
该研究发现,CD4+CD8αα+ IELs主要存在于无特定病原体(SPF)小鼠而不是无菌(GF)小鼠的小肠上皮内。因此,作者认为寄生于小肠的细菌通过修饰宿主饮食中的代谢产物来促进CD4+CD8αα+ IELs的诱导。随后,他们比较不同营养饮食的SPF小鼠肠道组织,发现营养丰富饮食的SPF小鼠CD4+CD8αα+ IELs水平显著高于较低限度营养饮食的SPF小鼠,饲粮对CD4+CD8αα+ IELs水平的影响仅限于小肠(十二指肠、空肠和回肠),在盲肠和结肠中未观察到。这表明,由宿主或其共生微生物修饰的饮食来源的分子,可能对小肠中CD4+CD8αα+ IELs的诱导至关重要。
接下来,作者试图确定饮食和肠道因素是否负责调节小肠中CD4+CD8αα+ IELs水平。在确定胆汁酸(BA)本身和BA信号级联不能调节小鼠小肠中CD4+CD8αα+ IELs水平后,他们最终将注意力集中在各种脂肪酸上。令人惊喜的是,在给与小鼠喂食特定脂肪酸后,他们发现,在食用亚油酸(LA)的小鼠小肠中出现了CD4+CD8αα+ IELs细胞。
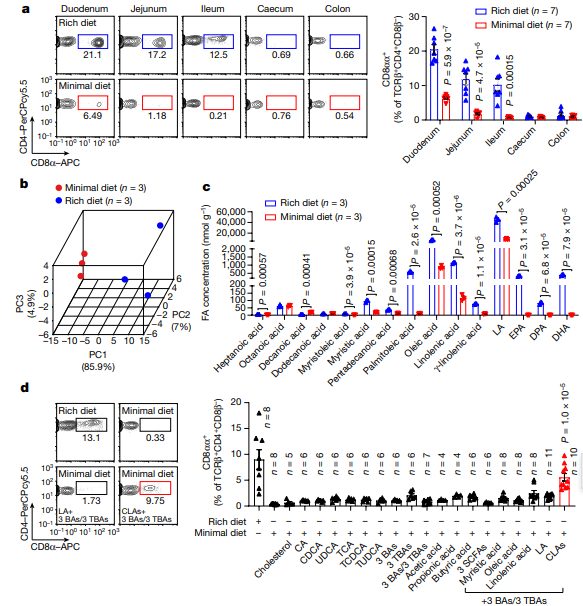
图1 肠道长链脂肪酸(LCFA)代谢物对上皮内CD4+CD8αα+ 细胞的维持至关重要
微生物是LA异构化的必要条件
一些肠道细菌,如肠球菌、乳杆菌、双歧杆菌、瘤胃球菌和毛螺菌,可以通过亚油酸异构酶(LAIs)将LA转化为CLAs。研究者发现摄入较低限度饮食小鼠的回肠中能够产生CLAs的菌群丰度降低,随后进一步确定饮食和微生物群对肠道中CLAs水平的影响,并发现细菌生物转化过程对肠道CLAs的产生至关重要。因此,他们推测携带LAI基因的肠道细菌可能是小肠中CD4+CD8αα+ IELs的诱导者。
为了进一步确定LAI基因阳性细菌中的LCFA异构化途径是否参与回肠CD4+CD8αα+ IELs的诱导,作者选择两种CD4+CD8αα+ IELs诱导株:粪肠球菌MMH594和粪肠球菌OG1RF进行探究。在失去将LA转化为CLAs的能力后,这些粪肠球菌菌株无法诱导回肠产生CD4+CD8αα+ IELs。这表明, LAI阳性共生菌编码的LA代谢途径是诱导小鼠小肠CD4+CD8αα+ IELs所必需的。
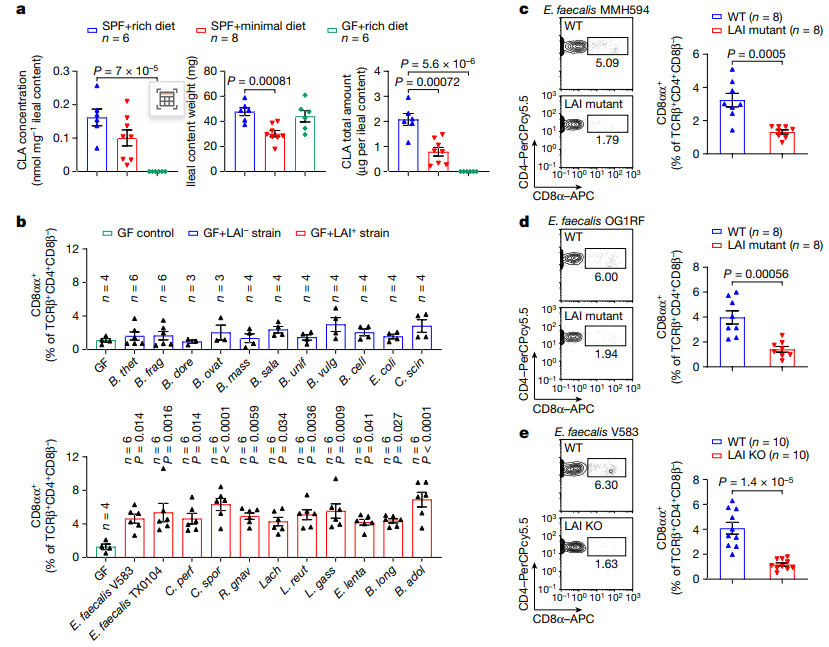
图2 肠道细菌通过LCFA异构化途径控制上皮CD4+CD8αα+细胞
CLAs通过HNF4γ诱导CD4+CD8αα+ IELs
饮食中的LA可以被微生物代谢修饰,其产物CLAs具有宿主免疫调节特性。因此,作者进一步探究宿主如何识别微生物CLA信号来维持回肠CD4+CD8αα+ IELs稳态。表达CD8α的SPF小鼠回肠IELs 转录组学分析显示,CD4+CD8αα+ IELs、TCRβ+CD8αβ+以及TCRβ+CD8αα+ IELs的基因表达模式明显不同。尽管所有三个IEL群体都表达了与CD8+ T细胞中细胞毒性T淋巴细胞程序相关的基因,但CD4+CD8αα+ IELs保留了许多由CD4+细胞系编码的细胞因子的基本表达,这些细胞因子(例如IL10、IL17a、IL17f和IL22)对维持肠道屏障功能非常重要。
进一步分析发现CD4+CD8αα+ IELs选择性地表达两种核受体: HNF4α和HNF4γ,这两种受体是肠道上皮细胞dFA代谢的关键调节因子。其中HNF4α在整个胃肠道中高度表达,而HNF4γ主要在小肠中表达。CD4+ T细胞中HNF4α的缺失并不影响回肠CD4+CD8αα+细胞或其他IEL对照物的水平。值得注意的是,TF-ThPOK和受损的CD4谱系水平升高HNF4g缺陷小鼠CD4+IELs中CD8αα+细胞频率。通过比较HNF4g缺陷小鼠及其对照组小鼠IECs中免疫信号相关基因的表达,发现这些免疫基因在Hnf4g缺陷的IECs中没有下调,这表明在Hnf4g缺陷小鼠中存在一种特殊的调节IELs的机制。
接下来,他们探索了HNF4γ是否通过过继形式将Hnf4g充足的或缺乏Hnf4g的初始CD 4+T细胞转移到Tcrb基因敲除小鼠体内来调节CD 4+T淋巴细胞内的CD4+CD8αα+ IELs。转移Hnf4g缺乏的原始CD 4+ T细胞的小鼠回肠CD4+CD8αα+ IEL表型与缺乏Hnf4g的小鼠或喂食少量食物的小鼠相似。这一结果表明,HNF4γ对回肠CD4+CD8αα+ IELs具有内在的调节作用。
为确定肠道CLAs是否通过HNF4γ调节回肠CD4+CD8αα+ IELs,研究者用六种胆汁酸(CA、CDCA、UDCA、TCA、TCDCA和TUDCA)混合乳化的CLAs治疗Hnf4g缺陷小鼠,发现CLA-BA混合物能够恢复低营养饮食的Hnf4g窝鼠的CD4+CD8αα+ IELs的回肠频率;而单独补充CLA无法恢复Hnf4g缺陷小鼠的回肠CD4+CD8αα+ IELs水平,这表明微生物CLA - Hnf4γ轴在调节这种IEL群体中起着关键作用。
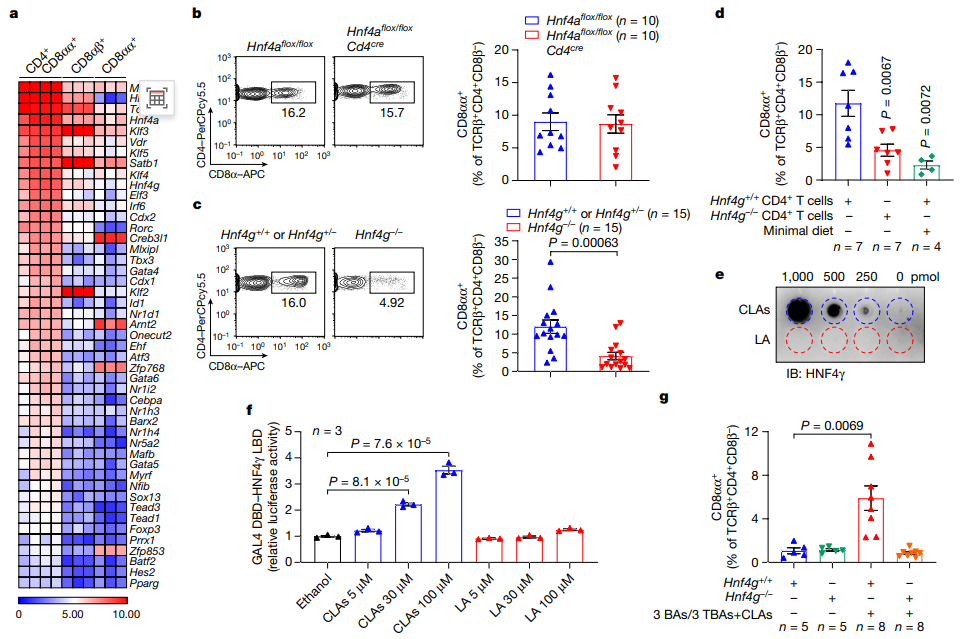
图3 肠道CLAs通过HNF4γ调节上皮内CD4+CD8αα+细胞
HNF4γ通过IL-18受体下调ThPOK
随后,研究者进一步探究CLA-HNF4γ轴调节CD4+CD8αα+ IELs的机制。三个表达CD8 α的TCRβ+ IEL亚群的转录组比较显示,CD4+CD8αα+ IELs高水平表达IL13ra1和IL18r1。检测相应细胞因子缺陷小鼠的回肠CD4+CD8αα+ IELs水平表明,IL-18信号传导在控制这种IEL群体中具有潜在作用。虽然HNF4γ缺乏或低营养饮食不会影响回肠IL-18水平,但HNF4γ可以结合回肠CD4+ IELs中IL18r1位点的转录起始位点(TSS)。
他们还发现低营养饮食导致CD4+ IEL表面IL-18R1表达受损。通过补充CLA,小鼠CD4+ IELs中IL-18R1水平的恢复依赖于HNF4γ。这表明CLA-HNF4γ轴在控制CD4+ IELs中IL18r1介导的信号传导中是至关重要的。进一步实验发现在IL18缺陷小鼠和IL18r1缺陷小鼠的CD4+ IELs中ThPOK+种群增加,而在这些小鼠的回肠中其他TCRβ+ IELs保持完整。因此,肠道CLAs激活HNF4γ促进IL-18R1在CD4+ IEL中的表达,而IL-18反过来通过其受体信号下调ThPOK,使CD4+CD8αα+ IELs分化(图4)。
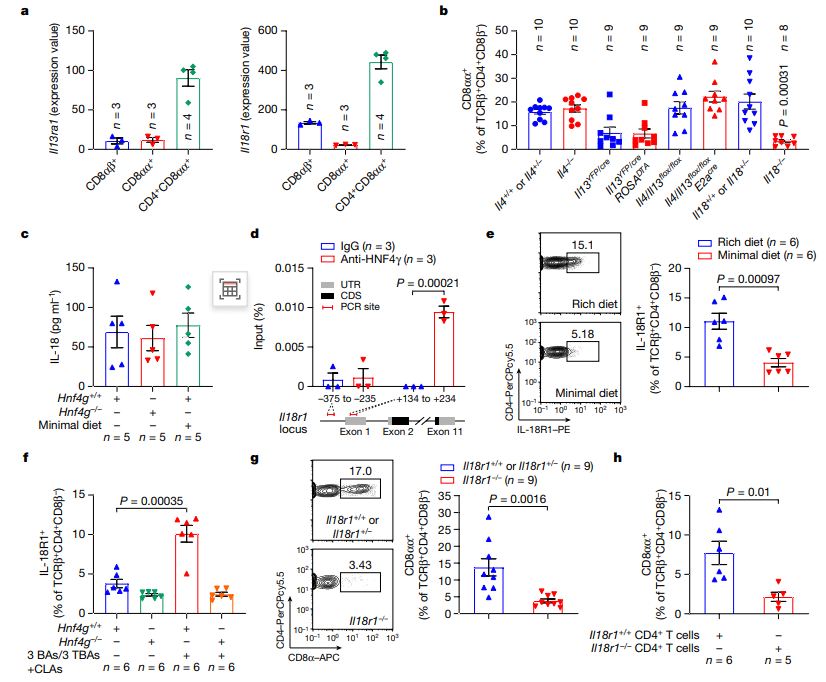
图4 肠道CLA-HNF4γ轴通过IL-18信号传导诱导上皮内CD4+CD8αα+细胞
CLA-HNF4γ轴调控肠道感染
CD4+CD8αα+ IELs已被报道可维持肠道内环境。因此,研究者分析了CLA-HNF4γ级联是否调节鼠伤寒沙门氏菌(Salmonella typhimurium,STm)感染模型中的肠道炎症反应。他们发现饲料中添加CLA可以恢复小鼠回肠的CD4+CD8αα+ IELs。此外,给予CLA+低营养饮食的小鼠对STm感染的抵抗力与营养丰富饮食的小鼠相同,而单纯喂食低营养饲粮的小鼠表现出更高的死亡率、严重的上皮破坏、血清炎症细胞因子水平增加以及更多细菌传播到非肠道器官等不良症状。因此,CD4+ T细胞中的HNF4α信号在STm感染的控制中是少不了的。然而,Hnf4g缺陷小鼠比同窝出生的小鼠更容易感染STm,CD4+ T细胞中的HNF4γ信号也显示出对STm感染的保护作用,表明干预CLA-HNF4γ轴可调控肠道感染程度。
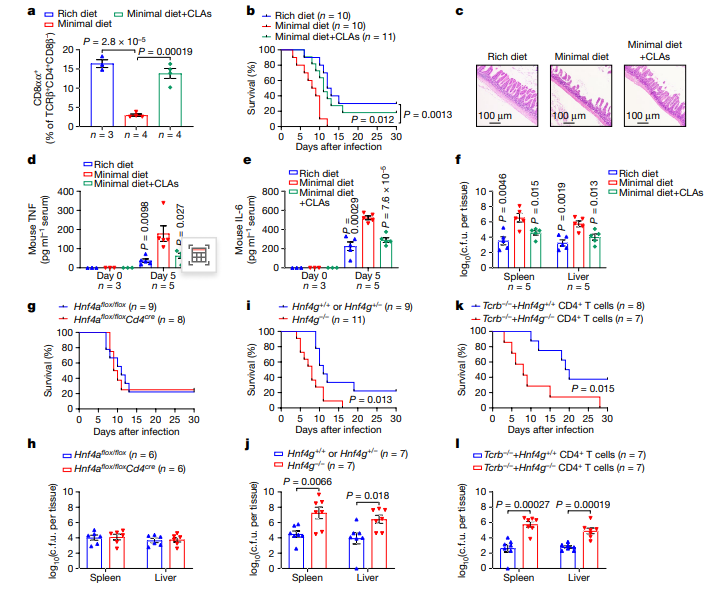
图5 CLA-HNF4γ轴控制肠粘膜感染
该研究通过靶向代谢分析结合体内外机制探索,表明肠道菌群FA衍生物CLAs是通过HNF4γ介导的IL-18受体信号传导维持健康的CD4+CD8αα+ IELs池的重要分子因子,提出了一种膳食-微生物-宿主的三重互作信号网络调控机体黏膜稳态与抗感染免疫的新型功能机制。
小爱科普:长链脂肪酸是一类复杂而又重要的膳食营养素,越来越多的研究揭示了其在肿瘤/癌症及肠道免疫等方面发挥的关键作用。上述研究由爱必信(上海)生物科技有限公司提供亚油酸同分异构体共轭亚油酸(abs47051161),除此之外爱必信还可为广大科研学者提供多种高品质生化试剂,包括脂类、酶类、植物提取物、荧光染料、氨基酸&多肽&蛋白质、碳水化合物、小分子激动剂/抑制剂等,助您科研探索之路不断创新和突破!
参考文献
[1] Song, X., Zhang, H., Zhang, Y. et al. Gut microbial fatty acid isomerization modulates intraepithelial T cells. Nature. 2023.
本期产品推荐:
货号 | 产品名称 | 规格 | 纯度 |
abs42024588 | 棕榈酸 | 500g | 98%,BR级 |
abs42024595 | 棕榈油酸 | 5g | ≥98% |
abs42027728 | 硬脂酸 | 5Kg | 98%,BR级 |
abs44103152 | 二十碳饱和脂肪酸 | 100mg | GC≥98%,标准品 |
abs44103153 | 花生四烯酸 | 20mg | GC≥99%,标准品 |
abs47000922 | 肉豆蔻酸 | 20mg | HPLC≥98% |
abs47001426 | α-亚麻酸 | 0.2mL | HPLC≥98% |
abs47001873 | 二十二碳六烯酸(DHA) | 50mg | GC≥98% |
abs47024476 | 亚油酸 | 50mg | HPLC≥98% |
abs47024483 | γ-亚麻酸 | 20mg | GC≥98% |
abs47047897 | 月桂酸 | 1g | GC>99.5%,标准品 |
abs47047898 | 月桂酸 | 500g | 98%,AR级 |
abs47051161 | 共轭亚油酸 | 2.5Kg | 异构体混合物,80% |
abs47051421 | 正癸酸 | 2.5L | AR,99% |
abs580141 | 游离脂肪酸检测试剂盒 | 96T | — |
* 温馨提示:absin所有产品仅用于科学研究,请勿药物、家用或其他用途。
好消息!Absin文献奖励重磅升级!

Absin特色产品线:






